Created by Kamila Klavíková
Kombinované zápisky z přednášek evoluční biologie, genetiky a evoluční genetiky na PřF UK. Vytvořené jako příprava ke státnicím.
evoluční model nukleotidových sekvencí
Předpoklady:
Nukleotidové sekvence se vyvijí rychleji. Jsou vhodné pro blízce příbuzné organismy.
Proteinové sekvence se vyvíjí pomalu. Hodí se tedy pro vzdáleně příbuzné skupiny. Více konzervované než nukleotidové sekvence. Různé kodony jsou více preferované pro danou aminokyselinu v různých druzích.
Ribozomální RNA se též vyvijí pomalu.
Modely mutací a evoluce jsou budované nad přiřazeními k odhadnutí evoluční vzdálenosti dvojice sekvencí.
Evoluční vzdálenost je odhadovaný počet mutací, který se objevil od doby, kdy se dvě sekvence oddělily od společného předka.
loss of heterozygosity (LOH)
Mutace alely určitého genu v somatické buňce, která již nese na homologickém chromozómu mutovanou alelu téhož genu.
Kanibalistická teorie. Jeden organismus sežral druhý organismus stejného druhu. Dostala se mu dovnitř cizí DNA. Vyvinula se mitóza. Diploidní organismy jsou odolnější. Zařadili cizí DNA do své.
Teorie promiskuitních bublin. Předbuněčné stádium. Silný horizontální genový přesun. Vyměňování genetických informací. Jejich zařazení. Meióza...
Teorie vzniku odolných diploidních stádií. Shlukování protoeukaryot za nepříznivých podmínek – možnost splývání.
Změna exprese, NE produktu; down mutace → snížení transkripce + zvětšení vzdálenosti od START nukleotidu; up mutace → zvýšení transkripce; zmenšení vzdálenosti od START nukleotidu.
V užším smyslu – nepříznivé podmínky na Zemi – důležitost oprav DNA – odstraňování mutací – když je jen jedna verze DNA jako předloha, vždy se bude chyba kopírovat – je nutné mít dvě, aby mohla být špatná sekvence opravena.
Reset epigenetických signálů a oprava ploidie. Není nejdůležitější rozdělit se, ale opravit se.
Dochází k omezení nežádoucí variability.
Netýkají se DNA; většinou při transkripci; projev obvykle slábne s časem.
Mutace v 3´sestřihovém místě → sestřihový aparát toto místo přeskočí a dojde k odstranění exonu z transkriptu
Mutace v 5´sestřihovém místě → zachován intron v transkriptu
Možný vznik eukaryot z konsorcia archea, protobakterií a velkých cytoplasmatických virů.
Vakcinační teorie. Genomoví parazité (např. viry) mohli ochraňovat před nebezpečnými viry.
Teorie nekódující DNA.
Teorie pásu cudnosti.
Somatické buňky jsou schopny dovršit pouze určitý, předem daný počet buněčných dělení.
Lidské fibroblasty – 50-60 mitóz
Vyjímku tvoří maligně transformované buňky.
Zygota tak má před sebou pouze omezený počet mitóz a pouze linie zárodečných buněk, které opakovaně procházejí pohlavními procesy, je vlastně „nesmrtelná“. Je možné, že po splynutí gamet dochází v zygotě k reparaci většiny mutací, ke kterým došlo v germinální linii během života mnohobuněčného organismu.
Somatické buňky takovým procesem neprocházejí. Zhoršuje se jejich funkčnost. Tkáně i celý organismus stárne.
K vysvětlení Hayflickova limitu je mnoho teorií, např. postupné zkracování telomer chromozomů během mitotických dělení a obnovení jejich původní délky po indukci enzymu telomerázy v některé fázi pohlavního rozmnožování.
Ve stejném místě jako byla mutace přímá.
Pravá reverze. Obnova mutované skevence nukleotidů na původní. Opačná reverze – změna kodonu na kodon synonymní k původnímu kodónu (před pímou mutací).
Částečná reverze. Změna kodónu na nesynonymní kodón, leč takový, že dojde k úplné či častečné obnově funkce s novou AK oproti mutaci přímé.
Vliv na expresi jednoho sousedního genu; geny lokalizovány na jedné straně mutantního genu.
Mutace v genu, který kóduje produkt účastnící se regulace transkripce jiného genu/mutace v regulační oblasti genu.
Lze využít pro stromy do 20 taxonů.
Absence pohlavního rozmnožování a s ním spojené reparace mohou být příčinou stárnutí klonů somatických buněk u mnohobuněčných organismů.
Nemožnost korekce téměř neutrálních mutací za nepřítomnosti genetické rekombinace může být příčinou stárnutí mnohobuněčných organismů.
Supresorová mutace. Maskuje účinek přímé mutace; intragenová (mutace v témže genu, ale na jiném místě) X intergenová (mutace v jiném genu než přímá mutace)
Enhancerová mutace. Opak supresorové; zvýšení intenzity mutačního fenotypu; často letalita.
Buněčný typ: somatické buňky / buňky zárodečné linie
Typ chromozomu: autozomální / gonozomální
Molekulární změna: substituce / inzerce / delece
Kořen umístíme doprostřed mezi dva nejvzdálenější taxony ve stromě. Předpokládá konstantní rychlost evoluce.
Outgroup je taxon, která je dost příbuzný na to, aby byla možná interfence s daty, ale dost vzdálený na to, aby byl jasný outgroup. Často určený z nezávislého zdroje informací. Př. outgrupu - ptačí sekvence u fylogenetické analýzy savců.
Rozmnožování patří mezi vlastnosti živých systému, bez nichž by nebyla biologická evoluce.
Pohlavní rozmnožování je až moderní akt této evoluce.
Dříve nejspíše sloužilo k jiným účelům, ale časem se na něj nabalila řada adaptací.
Velká část předností nemohla působit od začátku.
Rozdíl teorií vznik pohlavního rozmnožování a jeho udržování.
Přítomnost dvou genových sad pocházejících od dvou jedinců umožňuje zygotě rozpoznat, ve kterém z řetězců DNA došlo k mutaci.
Dojde-li k bodové mutaci, mohou enzymy reparačního aparátu určit místo, kde k této mutaci došlo. Báze v řetězci zde spolu nepárují. Nemohou ale určit, ve kterém řetězci došlo k nukleotidové zaměně. Je-li v buňce druhá kopie daného úseku DNA, místo ve kterém spolu báze nepárují, může být opraveno podle sekvence druhé kopie daného úseku DNA. Mohou být opravovány i neutrální mutace.
Přesto by tento mechanismus mohl být dlouhodobě nevýhodný, protože by danému druhu bránil v evolučním přizpůsobování se probíhajícím změnám. Je tedy možné, že v genomu by mohly být takto chráněny proti změnám pouze určité úseky genů.
Výhodnost schopnosti opravovat mutace v germinální linii je do určité míry sporná, ale schopnost opravovat mutace v liniích buněk somatických je jednoznačně výhodná.
Dominantní. Pokud projevena pozdě v ontogenezi, přenos do dalších generace.
Recesivní. V populaci zachovány hlavně u heterozygotů. Neobvyklé štěpné poměry v potomstvu F2 generace.
Auxotrofní. Neschopnost syntetizovat určitou důležitou živinu.
Teplotně senzitivní. Produkt labilní za určitých teplot.
Životaschopnost pouze za přítomnosti supresoru.
Ztrátová mutace. Snížení funkce (= leaky mutace) + ztráta funkce (= null mutace) mutovaného genu.
Zisková mutace. Zvýšení funkce; větší množství produktu; zisk nové vlastnosti
Počet nezakořeněných stromů je:
Somatické mutace. Projev u potomků somatické buňky; nepřenáší se gametami do potomstva; ztráta heterozygozity = mutace alely genu v somatické buňce, která už nese na homologickém CH mutovatou alelu téhož genu (retinoblastoma u člověka); dominantní mutace se projeví ve fenotypu
Gametické mutace. Přenos gametami do potomstva; dominantí mutace projev ve fenotypu potomstva
U většiny druhů množících se nepohlavně musí být série nepohlavních množení čas od času přerušená pohlavním množením. Jinak by došlo k postupné degeneraci populace. Jediný cyklus dokáže geneticky omladit danou populaci.
V průběhu pohlavního množení byly pravděpodobně nějakým zatím neznámým mechanismem opraveny mutace a poškození genetického materiálu.
Fylogenetika se zabývá studiem evoluční příbuznosti napříč různými skupinami organismů (např. druhy, populace).
Informace můžeme získat z fosilních záznamů, které obsahují informace o morfologii předků běžných druhů, avšak pro některé organismy nejsou dostupné (např. mikroorganismy...).
Dalším možným zdrojem jsou geny. V průběhu času shromažďují mutace způsobující fenotypové změny. Je pro ně dostupné větší množství dat.
Předpoklady:
U kladogramu větve nenesou žádnou informaci o evoluci (nekorespondují s množstvím změn v genech).
U fylogramu větvě představují množství evoluční divergence.
Formát, který je reprezentovaný vnořenými závorkami. Na fylogramu založený formát obsahuje délky větví.
Také známý jako hypotéza ostrých loktů.
Předpokládá, že polymorfismus potomstva omezuje konkurenci mezi sourozenci. Tím, že se sourozenci od sebe liší, mají i odlišné ekologické nároky (např. potravní).
Bodové mutace. Záměna dvou nukleotidů (inzerce, delece, transverze – různý typ báze, tranzice – stejný typ báze)
Řetězcové mutace. Záměna celého úseku DNA (duplikace, inzerce = zmnožení řetězce DNA, delece = vystřihnutí úseku DNA, translokace = vzájemné vyměnéní úseku DNA, inverze = určitý úsek DNA je z chromozomu vystřihnut a vložen do stejného místa v opačné orientaci – může způsobit reprodukční bariéru a tím speciaci)
Chromozomální mutace. Translokace velkého rozsahu vznikající při mitóze. Následky drastické – mládě se ani nenarodí, nebo nemoci typu Downův syndrom.
Genomové mutace. Probíhají na úrovni chromozomů či chromozomových sad v důsledkem poruch v průběhu buněčného dělení (aneuploidie, polyploidie, autopolyploidizace, aloploidizace)
Fylogenetický strom ukazuje evoluční vztahy mezi různými taxony, u nichž se věří, že mají společného předka.
Aby mohl být určitý znak předmětem přirozeného výběru není nutné, aby přímo zvyšovaly životaschopnost či plodnost svého nositele.
Stačí, když zvýší šance, že blízcí příbuzní (nejčastěji potomci a sourozenci) se dožijí reprodukčního věku. Jedná se o vlastnost zvyšující inkluzivní zdatnost jedince.
U tohoto znaku se uplatňuje příbuzenský výběr (kin selection). Příbuzenský výběr se ve svém mechanismu blíží výběru individuálnímu.
Vychází ze skutečnosti, že areál výskytu kteréhokoli druhu je většinou v té či oné míře heterogenní. Na každé mikrostanoviště se přitom dostane v každé generaci víceméně náhodný vzorek potomstva různých rodičů. Jedinci si mezi sebou na stanovišti konkurují a pouze nejlepší z nich v konkurenci zvítězí a zanechají potomstvo, tj. jejich fenotyp nejlépe odpovídá vlastnostem daného mikrostanoviště.
změny smyslu (vlivu na translaci)
Synonymní mutace. Substituce v jedné bázi, která nemění kódovanou AK.
Missense mutace. S chybným smyslem. Změna kodonu pro jednu AK za kodon pro jinou AK.
Nonsense mutace. Nesmyslná substituce vedoucí ke vzniku stop kodonu na místě kodonu pro AK.
Indukované mutace sou způsobené mutageny, což jsou látky, faktory vnějšího prostředí, které indukují vznik mutací. Jsou různé povahy, biologické, fyzikální a chemické.
Pohlavní rozmnožování.
Amfimixis, tj. střídání redukčního dělení meiózy s obnovením diploidie zárodečné buňky zygoty syngamií, tj. splynutím jaderného materiálů dvou haploidních pohlavních buněk pocházejících zpravidla od dvou jedinců stejného druhu.
Nepohlavní rozmnožování.
Automixis. Rozmnožování zahrnující obnovení diploidie splynutím dvou haploidních produktů meiózy stejného jedince.
Apomixis. Rozmnožování nezahrnující meiózu.
Určitá aplikace evolučního principu Červené královny. Princip pojmenovaný podle knihy Lewise Carrolla.
Říká, že v některých situacích je nutné běžet rychle kupředu, abychom zůstali aspoň na místě. Abychom se pohnuli vpřed, nestačí jen běžet, je nutné běžet rychleji než ostatní.
Hypotéza uvažuje skutečnost, že v prostředí, v němž za většinu selekčního tlaku jsou zodpovědné především biotické faktory, především paraziti a predátoři, je mnohdy výhodné se lišit od vlastních rodičů a od většiny dalších jedinců stejného druhu.
Extrémní tlak tohoto druhu vykonávají právě parazitické organismy, bakterie, viry... Populace hostitelského organismu může být zdecimována svým parazitem. Přežijí jen ti rezistentní, kteří dají vzniknout nové populaci jedinců. Parazit má ale v koevolučních zápase neustále náskok (kratší generační doba oproti hostiteli, větší selekční tlak...). Jediná účinná protistrategie hostitele je produkce různorodého potomstva. Odolnost proti epidemii přitom vykazuje výrazně negativní dědičnost.
Část neutrálních mutací je slabě negativní. V obrovské populaci by se chovaly negativně.
Počet mutací v dané pozici za jednotku času všech členů populace.
Výskyt dané mutace vyjádřen jako podíl mutantních jedinců v populaci. Vyšší frekvence mutací u samců (vyšší selekční tlak na X chromozom a vyšší počet buněčných dělení během spermatogeneze).
Spontánní mutace vznikají přirozeně bez známé příčiny.
Při deaminaci dochází k nahrazení aminoskupiny (-NH2) ketoskupinou.
Cytosin. Hydrolýza na uracil.
5-methyl-cytosin. Hydralýza na thymin. Místo nejčastějších mutací u obratlovců.
Adenin. Vzniká hypoxantin, který se raději váže na cytosin. Vzniká tedy v podstatě GC pár.
Guanin. Vzniká xantin, který se páruje s thyminem. Nový pár je v podstatě AT.
Ztráta purinové / pyrimidinové báze a vznik apurinického / apyrimidinového místa (AP místa).
Př: ztráta purinové báze (A, G) ⇐ přerušení glykozidické vazby mezi deoxyribózou a purinem
Sklouznutí (slippage) je sklouznutí DNA polymerázy během replikace, lokální denaturace a následná reparace. Dopředné sklouznutí – delece, zpětné sklouznutí – adice. Může k němu dojít kdekoliv v DNA. Nejčastěji se tak děje v repetetivních sekvencích.
Báze je pozměněná změnou pozice vodíkového atomu. Pozmění se způsob tvorby vodíkových můstků, čímž se změní párování. Stabilní formy bazí – keto (G, T) a amino (A, C), méně stabilní enol a imino.
Pohlavní rozmnožování úzce souvisí se vznikem fenoménu biologického druhu. Ke vzniku mohlo dojít až se vznikem pohlavního rozmnožování.
Nepohlavně se množící organismy. Druhy v pravém slova smyslu neexistují. Organismy často vytváří jakousi spleť více či méně si podobných a více či méně si příbuzných genealogických linií.
Např. pampeliška.
Pohlavní rozmnožování. Je s ním spojené neustálé promíchávání genů v rámci společného genofondu. Je to nejdůležitější mechanismus druhové koheze, což je mechanismus udržující po dlouhé období vlastnosti příslušníků jednotlivých druhů v určitém úzce vymezeném rámci.
Mnoho nevýhod:
Klasický darwinismus je založen na postupné kumulaci mikromutací, tedy mutací, které způsobují malé změny oproti původnímu stavu.
Existují ale i makromutace, jejichž vlivem se stavba či fungování těla změní výrazným způsobem (třetí ruka, jedna noha...). Je však otázka, zda by takovýmto způsobem mohly vzniknout některé zásadní nové orgány jako např. křídla apod. Makromutace existují, ale jejich role v evoluci je nejistá. Jedině díky makromutacím můžou vzniknout mezidruhové rozdíly.
Mutace jsou změny ve struktuře genetického materiálu, při nichž se mění smysl genetické informace, aniž by byla porušena syntaktická pravidla jejího zápisu. Jako mutace označujeme změny sekvence DNA.
Je třeba rozlišovat mutace, respektující pravidla zápisu do DNA, a poškození DNA. Mutace jsou největším zdrojem genetické variability.
Genové mutace. Postihují jen jeden gen.
Chromozomové mutace. Změna struktury chromozomů, pořadí genů, polohy centromer.
Genomové mutace. Celková změna genomu (polyploidizace, aneuploidizace).
Místa výskytu mutací nejsou v řetězci DNA rozmístěna rovnoměrně – častěji k nim dochází na horkých místech (hot spots). Změny frekvence mutací pozorujeme i v čase, kdy jich v některých místech evoluce vzniká podstatně více – není úplně objasněno proč – sekvenční motiv (AT sekvenční motivy), vnější podmínky, pohlavní rozdíly (samci mutují častěji)... Jsou také místa, kde sice nevzniká více mutací, ale kde se mutace snáze fixují.
Některé organismy dokážou ve stresových podmínkách začít mutovat rychleji, což jim zvýší šanci na přežití. Někteří autoři se domnívají, že nejen mutují rychleji, ale i cíleně, např. trypanosoma dokáže vytvářet mnoho různých variant povrchových proteinů a dělá to záměrně v určitý čas a za určitých podmínek.
Liší se mezi pohlavími. Častěji mutují samci – při vzniku spermií mohou zkoušet evoluční novinky. Samice jsou konzervativnější, má jich méně a nevyplatilo by se jí riskovat.
Role mutátorů – patogenní bakterie mají v populaci hodně mutátorů, snaží se tak najít optimální virulenci.
Částečně zanesené evoluční info o potenciálně odlišné rychlosti změn v různých větvích.
Na začátku strom - hvězda.
Vypočíst matici vzdáleností pro všechny dvojice sekvencí. Modifikovat vzdálenosti kvůli kompenzování dlouhých hran.
Vybrat minimální vzdálenost Di,j.
Definovat nový vrchol k a nastavit vzdálenost dkm. Přidat k do stromu.
Myšlenka možnosti přechodu jednoho druhu v jiný prostřednictvím přepínání alternativních ontogenetických programů. Př. když se pšenici nedaří, začne se měnit na žito, kterému by se za daných podmínek dařilo lépe.
Tato myšlenka není úplně zavrhnutelná, ale v rámci celého lysenkismu (falšování, ideologie) je mimo.
Kvantitativní měřítka jako vzdálenosti a podobnosti mezi druhy.
Nic neříkají o stavu předků ve vnitřních uzlech. Spoustu informací jsme ztratili převodem na matici vzdáleností.
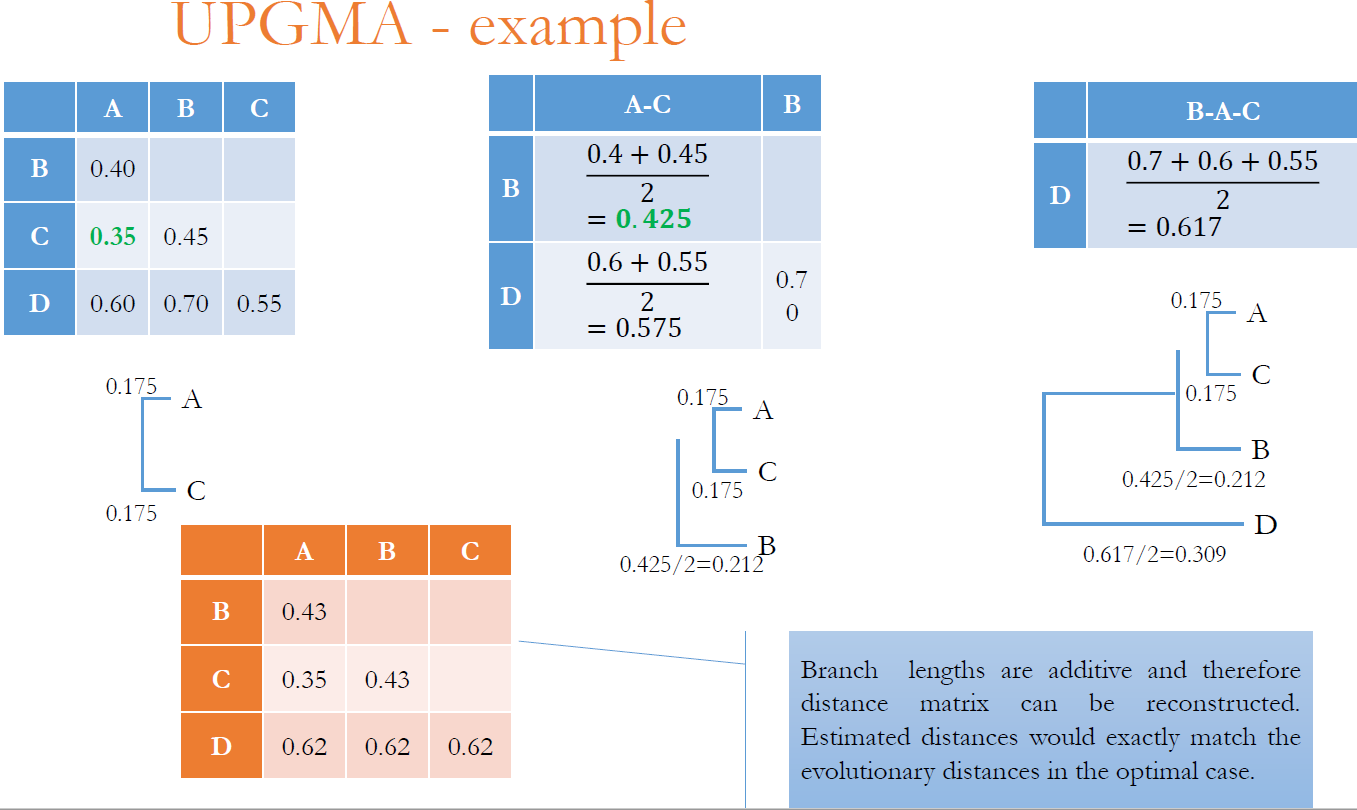
Unweighted Pair Group method Using Arithmetic Average
Předpoklad: molekulární hodiny – všechny taxony se vyvíjí konstantní rychostí a všechny jsou stejně vzdálené od kořene.
Metoda je založená na klastrování sekvencí:
Předpokládá molekulární hodiny, kde se předpokládá, že divergence sekvence se odehrává stejnou rychlostí kdekoliv ve stromu. Součet časů podel každé větvě je stejný. Pokud je to pravda, jedná se o správně vytvořený strom. Nebere v úvahu různé substituční rychlosti na různých větvích.
Je validní pouze pro ultrametrické vzdálenosti. Pro každou trojici sekvencí si, sj, sk jsou vzdálenosti dij, djk, dik buď všechny stejné, nebo dvě vzdálenosti jsou stejné a zbývající je menší. V ultrametrickém stromu jsou vzdálenosti z kořene do listů konstantní.
Druh je skupina populací, mezi kterými může dojít ke křížení a které jsou reprodukčně izolovány od jiných populací (E. Mayr).
Není úplně pravda.
Samostatná existence cílených mutací by ale nestačila k lamarckistické evoluci. Naprostá většina mutací je náhodná.
První překážkou lamarckistické evoluce je neexistence zpětného toku z proteinů do DNA.
Druhou překážkou je Weismanovská bariéra mezi germinální a somatickou linií.
Třetí překážkou je fakt, že geneticka informace není přesný popis struktury.
maximum likelihood -> pravděpodobnost dat ze stromu (modelu)
BT -> pravděpodobnost stromu/uzlu při datech (využívá k výpočtu tzv. inverzní pravděpodobnost)
Využívá komplexní samplovací metody (př. Markovské řetězce Monte Carlo).
Nejméně anagenetických změn, nejpravděpodobnější průběh kladogeneze.
Metoda maximální parsimonie se snaží najít takový strom, kde bude, co nejméně změn.
Algoritmus má 2 části:
Jean-Baptiste Lamarck
před Darwinem (před r. 1859)
Př. žirafy mají dlouhý krk, protože se natahují pro listí do korun stromů.
Pravděpodobnost pozorování našich dat za předpokladu, že platí náš model.
model = strom + parametry
Pravděpodobnost poskytuje pravděpodobnosti sekvencí pro daný model jejich evoluce v určitém stromě. Čím pravděpodobnější jsou sekvence pro daný strom, tím více je strom preferovaný. Berou se v úvahu všechny stromy.
Paralela k fylogenezi: máme model (strom + parametry evoluce) a zkoumáme jaká je pravděpodobnost, že na ně budou sedět naše data, testujeme postupně stromy (analýza tedy vezme 1 strom, spočítá L, pak vezme další, a pokud ten je lepší tak pokračuje s ním, pokud ne tak s původním) – počet kroků = počet generací -> vyhledáme tímto nejlepší strom.
Příklad s házením mincí: hodím 10x mincí, jaká je pravděpodobnost (likelihood, L), že mi padne 5 x hlava a 5 x orel? (náš strom je jakoby to, co „má“ padnout, parametry jsou třeba pravděpodobnost, že padne hlava, kolikrát jsme házeli apod.)
Pokud by mince byla nesouměrná, a měnila by se pravděpodobnost (p) padnutí hlavy, pak se zmenšuje i likelihood padnutí 5 hlav + 5 orlů
Homologie je znak zděděný od společného předka.
Př. končetiny obratlovců.
Analogie je jen podobný znak u nepříbuzných organismů.
Př. křídla u netopýrů a ptáků, ploutve u ryb a kytovců.
U druhů s nepohlavním rozmnožováním je to jednodušší. Když dojde k mutaci, může přílušná linie dát vzniknout novému druhu. Zda vznikne závisí na tom, zda existuje v prostředí pro takový druh volná nika, nebo zda je jeho fitness podstatně vyšší než původní varianty.
Pohlavně se rozmnožující druhy to mají složitější. Nestačí odlišné fenotypy a niky, musí také vzniknout reprodukční bariéra. Aby mohlo dojít alespoň k fyletické speciaci, je nutné, aby nová mutace fungovala i v ostatních kombinacích genů než jen v mutantovi.
Změna fitness za generaci je rovna variabilitě v populaci.
biologická zdatnost
Kvantitativní reprezentace přirozeného a pohlavího výběru. Lze ji definovat různě.
Může ji posuzovat např. podle toho, kolik daný jedinec zanechá potomků. Lepší je počet prapotomků nebo praprapotomků. Je to na hovno, pokud jsou to chcípačci, kteří se stejně nerozmnoží.
Biologickou zdatnost lze vyjádřit absolutně i relativně, kdy jedinec s největším počtem potomků má fitness = 1.
Fyletická speciace, kdy se postupně mění jeden druh jako celek v jiný druh (mění se fenotypové vlastnosti jeho příslušníků).
Při štěpné speciaci se jeden druh mateřský rozpadne na dva či více druhů dceřiných, které se co do svých fenotypových vlastností vyvijí samostatně.
Počítání počtu substitucí.
Algoritmus vypočte minimální cenu Rk(a) přiřazení písmene a do vrcholu k v místě u.
Cena S(a, b) pro každou substituci ve stromě.
Používá se pro rekonstrukci kladogeneze, protože když dva druhy sdílí hodně pleziomorfií, nemusí si být příliš příbuzní, ale mohou se jen málo měnit a žít třeba ve stejném prostředí. Ale když sdílí hodně synapomorfií, mají asi společného předka, který není předkem ostatních studovaných druhů
Jedním ze směrů popírajících darwinismus je mutacionismus. Jeho zastánci tvrdili, že nejdůležitějším zdrojem mutací jsou reparační procesy. Nebyli tak ale schopni objasnit vývoj adaptivních znaků. To vysvětluje mechanismus přirozeného výběru, který je nepostradatelný pro šíření výhodných mutací.
Kladogeneze je postupné odštěpování evolučních linií od linie vedoucí ke společnému předku.
Pleziomorfie je původnější forma znaku v dané linii.
Např. pětiprsté končetiny u tetrapodů, zuby u všech tetrapodů kromě ptáků a želv.
Apomorfie je odvozenější forma, přítomnost evoluční novinky.
Např. křídla ptáků, bezzubost ptáků a želv.
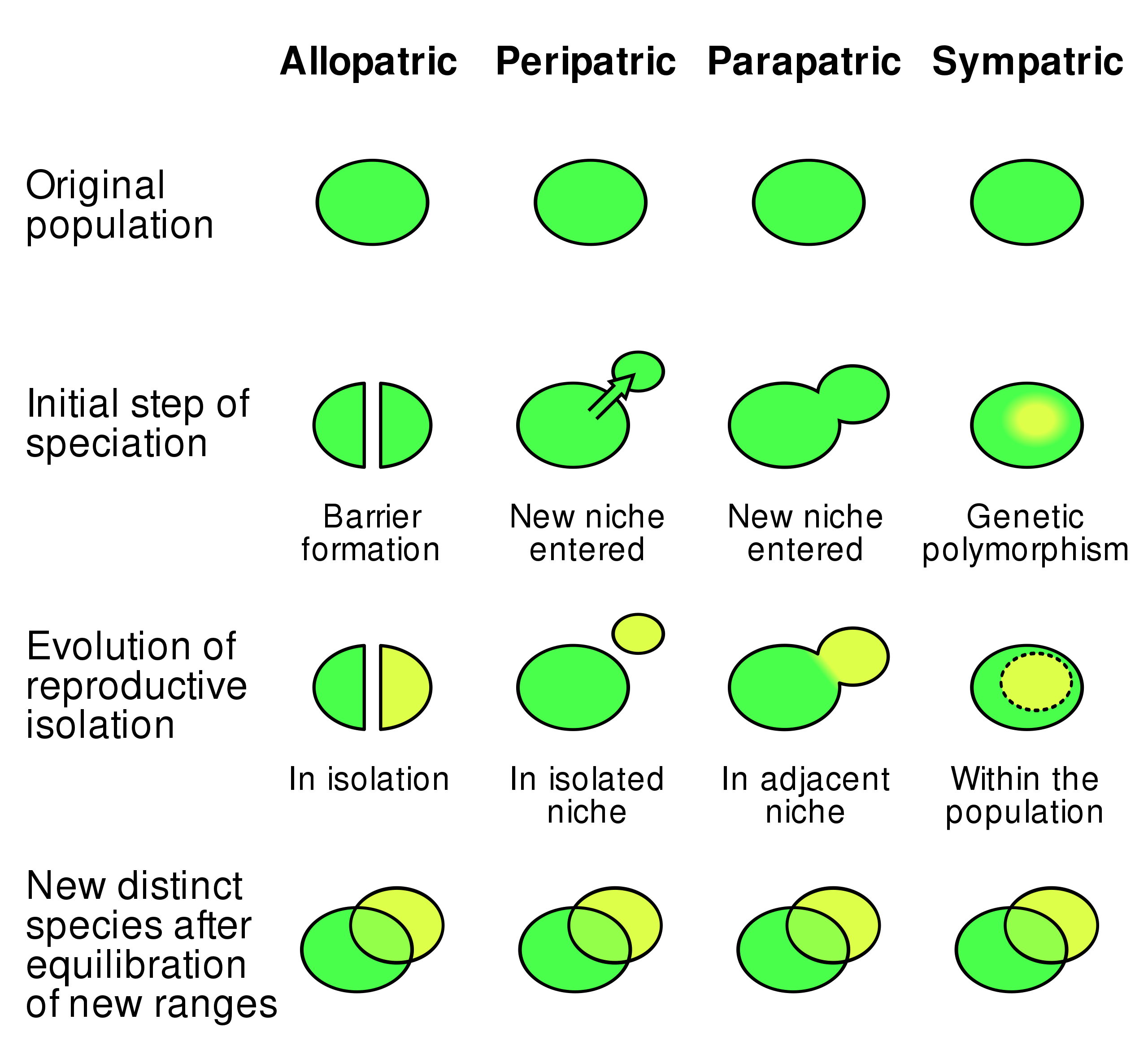
Speciace je proces, kdy vzniká z jednoho druhu jeden či více druhů nových.
Charles Darwin
shrnuto v jeho knize „O původu druhů přirozeným výběrem“
Postupné speciace, kam patří většina speciací, trvají dlouhou dobu (dloudo trvá vytvoření reprodukčně izolačních bariér). Př. alopatrická speciace
Okamžité speciace, které mohou proběhnout téměř v jeden okamžik. Př. speciace polyploidizační – v rámci 2n druhu vznikne 4n jedinec v důsledku poruch b. dělení, který se fenotypově liší od svých předku, speciace extinkční -
studium vzorců chování a mechanismů jejich evolučního vzniku
chromozomová speciace – při chromozomové mutaci se změní morfologie nebo počty chromozomů. Starý a nový karyotyp pak nemusí být kompatibilní. Otázkou je, zda to vytváří RIM, nebo se toto vytvoří až po oddělení druhů. Některé typy chromozomových speciací předpokládají spoluúčast většího počtu chromozomových mutací.
polyploidizační speciace – příčinou speciace může být genomová mutace
hybridizační speciace – rekombinační speciace / polyploidizace hybrida
ekologická speciace – u sympatrie, dvě formy druhu se izolují vytvořením prezygotických bariér
etologická speciace – změna v druhově specifickém rozpoznávání partnerů
vikariantní
Původně jednotná populace je rozdělená nějakou bariérou (řeka, pohoří, ...) na dvě srovnatelně velké populace, které se časem geneticky i fenotypově rozrůzní.
Williams a Hamilton
Richard Dawkins zpopularizoval (Sobecký gen)p
Při alopatrické speciaci nový druh vzniká postupně mimo kontakt s původní populací. Reprodukční bariéry se tvoří v oddělení.
Evoluce druhů má nespojitý charakter a probíhá ve skocích
Eldredge a Gould
vysvětluje se tím absence mezičlánků
punktuacionalistická
Během sympatrické speciace se nový druh formuje na stejném území, na kterém se vyskytuje druh mateřský. Buď došlo k diferenciaci nik, nebo může jeden z druhů druhý druh lokálně či globálně vytlačit.
Dobře je doložena pouze u cichlid v kráterových jezerech.
Začátek jako alopatrická, potom ale zmizí bariéra a dochází k sekundárnímu kontaktu obou vzniklých populací.
Nalezneme ji např. u nepříliš pohyblivých druhů, jejichž příslušníci vytvářejí v rámci svého areálu rozšíření vzájemně sousedící lokální populace.
Tok genů mezi populacemi je omezený, a je zde tendence k odstraňování hybridů vznikajících na „hranici“, čímž se tok genů ještě více omezuje.
Selekce proti hybridům, areál druhu, který se liší prostředím – př. část les a část louka, druh se postupně adaptuje na odlišné části areálu a časem vznikne RIM.
Z mateřské populace se oddělí jen velmi malá subpopulace, která dá posléze vzniknout novému druhu.
Je častější než dichopatrická, protože oddělení malé subpopulace je snadnější. Na druhou stranu většina těchto subpopulací zanikne, nebo opět splyne s hlavní populací.
Druhy žijí na stejném území. Jejich rozšíření je ostrůvkovité.
Př. parazit přenášený přímým kontaktem náhodně přeskočí na jiný druh hostitele, který žije na stejném území jako původní hostitel.
A ring species is a situation in which two populations which do not interbreed are living in the same region and connected by a geographic ring of populations that can interbreed.
Př. racci okolo polárního kruhu.
Evoluční soupeření probíhá na úrovni alel.
Richard Dawkins – model modrovousů.
U pohlavně se rozmnožujících organismů se fitness nedědí, je dáno genotypem (fenotypem), který se vytváří u každé generace de novo. Alely se ale předávají v nezměněné formě. Alela, která naprogramuje svůj organismus tak, aby byl co nejúspěšnější bude do další generace předána ve více kopiích než jiné alely téhož loku. Modrouvousovské geny jsou velmi rozšířené a úspěšné, nepřeváží proto, že populace, která je nese neuspěje v kompetici.
Motó Kimura
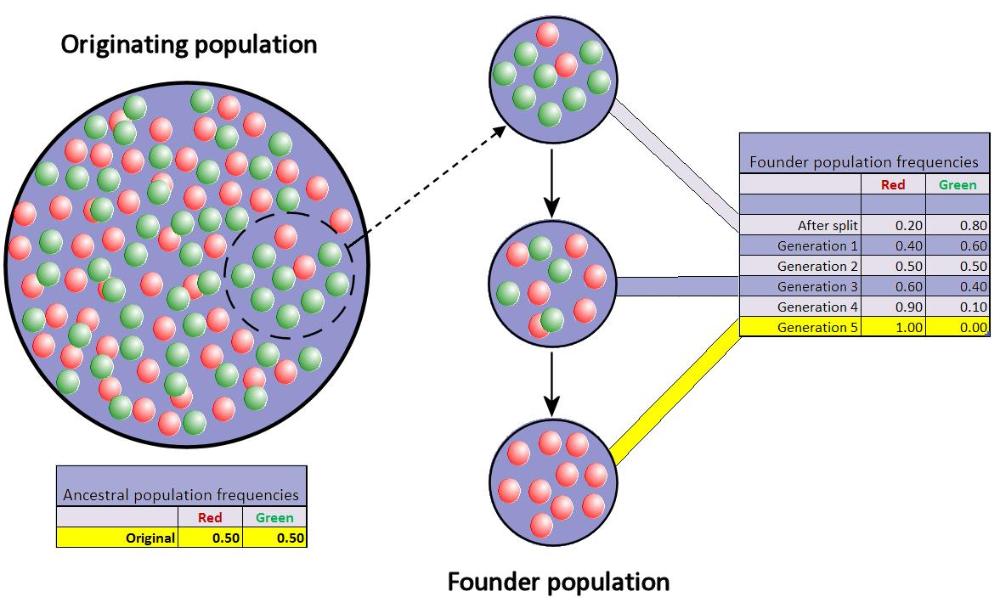
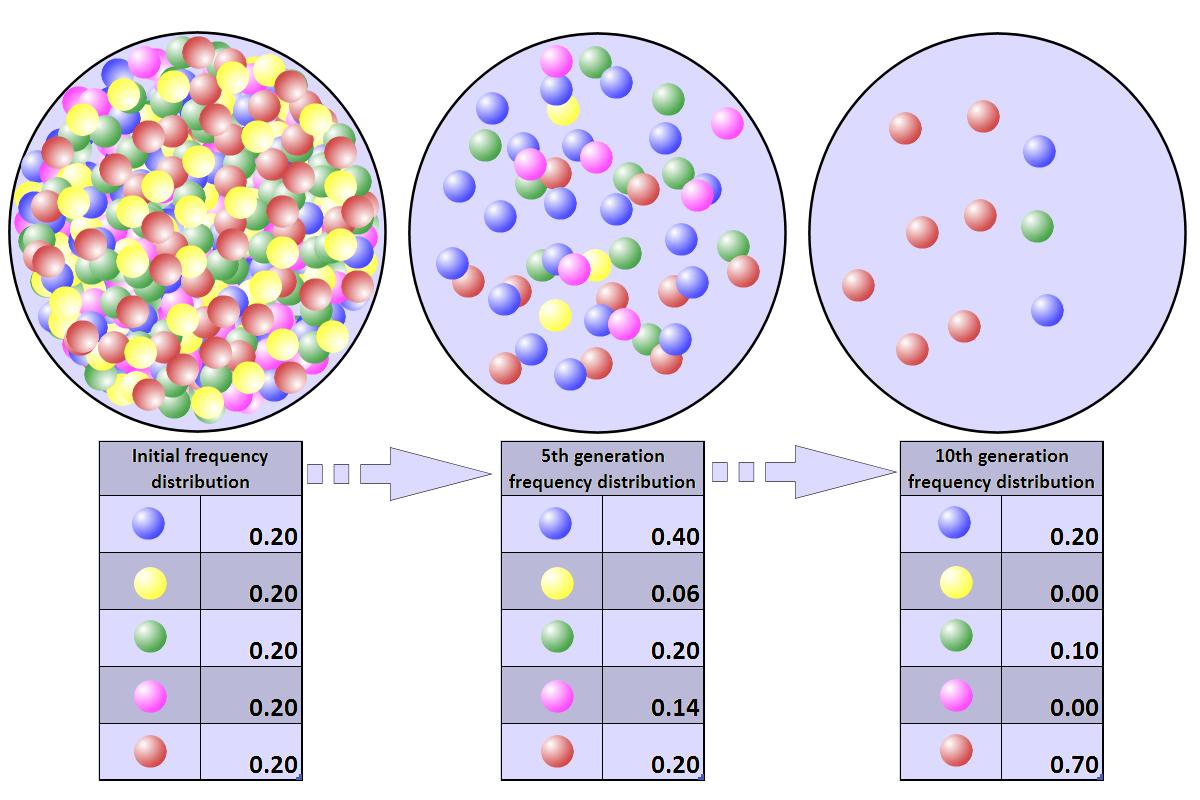
Při vzniku biodiverzity může mít zásadní vliv genetický drift.
Malá skupina v populaci se oddělí z původní populace a vytvoří novou populaci. Náhodný vzorek alel.
Zdatnost nosičů určité alely závisí na jejich četnosti v populaci. Jde o jeden z nejdůležitějších mechanismů pro udržení polymorfismu v populaci.
Pozitivní – konformita samic – Samice si vybírají podle toho, koho si vybere nejvíce samic. Předají to svým potomkům. Ti budou opět žádání. – včely se naučí létat na žluté kytky, proto je výhodné být žlutá.
Negativní – vstavači – 2 formy vstavačů, které nemají opylovačům, co nabídnout, jsou stejně zastoupené. Pokud bude jedné formy více, čmeláci se na ní naučí nelétat a bude to nevýhodné (červený). – dravec a kořist – Dravec se přednostně zaměří na nejpočetnější kořist. Postupně se její stav sníží a nejvýhodnější bude jiná kořist.
individuální výběr. Je výhodný pro jedince. Bývá nejsilnější. Ve většině případů silnější než skupinový. Zabýval se jím Darwin a je stále považovaný za nejdůležitější. Např. sexualita je vlastnost vzniklá tímto výběrem.
skupinový výběr. Vlastnosti výhodné pro skupinu, ale ne vždy pro jedince. Soupeří mezi sebou populace. Přítomen tam, kde se vytváří velké množství samostatných sociálních skupin stád, tlup, hejn.
příbuzenský výběr. Nelze jej zaměňovat se skupinovým. Existence altruistického chování, kdy jedinec snižuje své fitness ve prospěch celku. Určené primárně na pomoc blízkým příbuzným. Jde o zvýšení inkluzivní zdatnosti. Soupeři mezi sebou rody. Sobci v altruistických skupinách nikdy nevymizí, ale zároveň se jejich počet ani nezvyšuje.
druhový výběr. jednotlivé druhy spolu soupeří o to, který bude odštěpovat více dceřiných druhů, a o to, který bude s menší pravděpodobností podléhat extinkci.
mezidruhový výběr. pokud je znak, který je pro jedince neutrální a druhu prospívá, může se tento znak prosadit. Soutěží se o lepší využití zdrojů.
celospolečenstvový výběr. dosti pochybné, že by to mohl být předmět evoluce.
Znaky výhodné pro vyšší jednotku, ale neutrální pro jedince se mohou fixovat na vyšších úrovních.
Pseudoaltruistické chování jedinců stejného biologického klonu v eusociálních společenstvech je produktem individuálního, nikoliv skupinového výběru.
Během krátké doby dojde k razatnímu zmenšení populace. Kdo přežije je náhoda.
Intersexuální selekce je dána samičí volbou. Je zde rozdíl v množství energie vložené do vychování mláďat.
Samci mohou zvyšovat počet svých potomků zvýšením počtu sexuálních partnerek.
Samice mohou zvyšovat kvalitu potomstva výběrem samce.
Druhotné pohlavní znaky plní funkci indikátorů kvality jedince.
Znaky nefalšovaně vypovídající o zdatnosti samce (kdyby se brzy fixoval u všech, pohl. výběr by skončil), proto samci mají opatření proti podvodům.
Preferuje samce, kteří se dokáží prosadit proti ostatním samcům. K tomu mají vyvinuté druhotné pohlavní znaky.
Je dána několika faktory:
genetický posun
Genetický drift jsou změny frekvence alel. Náhodný posun alel. Některé alely se „vytáhnou“ častěji.
Toto kolísání je nápadné zejména v malých populacích. Může dojít k fixaci nebo ztrátě alely. Pokud bude populace dostatečně velká, nezafixuje se to nikdy. Čím větší populace, tím menší efekt driftu.
Jsou i jiné příčiny změny frekvencí jako selekce, mutace, migrace. Neuvažujeme tu selekci.
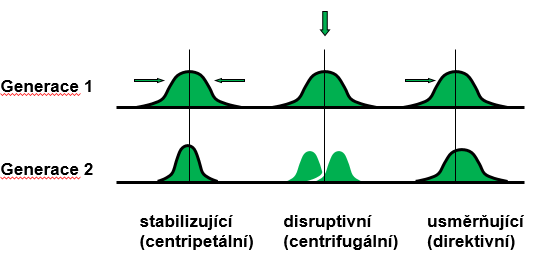
Stabilizující selekce odstraňuje jedince s extrémními hodnotami znaku.
Disruptivní selekce odstraňuje jedince s průměrnými hodnotami a zvýhodňuje extrémy.
Usměrňující selekce je posun maxima určitým směrem, obvykle jen na přechodnou dobu - reakce na změnu podmínek.
Veškeré typy výběru mohou existovat ve 2 zákl. formách:
přirozený výběr
Přirozený výběr je proces nerovnoměrného předávání alel pocházejících od jednotlivých individuí do genofondu následujících generací prostřednictím potomků.
Zahrnuje minimálně výběr přírodní a výběr pohlavní.
Podmínky fungování selekce:
Působením pohlavního výběru jsou selektováni jedinci s větší sexuální zdatností.
Vede k rozdílné evoluci morfologických znaků u obou pohlaví, ke vzniku druhotných pohlavních znaků (epigamních znaků), a tedy často i velmi nápadného pohlavního dimorfismu.
Pohlavní výběr mezi jedinci jednoho druhu, obvykle samci. Nestačí jen přežít, je třeba se více rozmnožit.
Jsou dva způsoby, jak toho dosáhnout:
Vysvětluje existenci excesivních znaků (př. páv), které snižují životaschopnost svým nositelům, ale samečka to zvýhodňuje.
Negativní selekce - odstraní se jedinci bez požadované vlastnosti.
Pozitivní selekce - podpora jedinců s požadovanou vlastností.
Genový tok je předávání genů mezi populacemi, nejčastěji pomoci migrujících jedinců.
V rámci populace je patrně nejdůležitějším zdrojem evolučních novinek (již odzkoušené) a napomáhá udržet genetický polymorfismus populace.
Protiváha genetického driftu. Stačí 1-2 migranti.
Přírodní výběr selektuje jedince s vyšší plodností a životaschopností.
Pokud jsou dvě alely současně pozitivně selektované, rychlost jejich šíření je menší než kdyby se šířily samostatně.
Pro fixaci alely je třeba, aby vymřeli jedinci bez této alely. Pokud ale mají jinou pozitivní alelu, vymírají pomaleji.
V komplexních systémech rozhodují o osudech nových mutací především evolučně stabilní strategie. Nemůže být převálcována jinou strategií.
Strategie jestřab (popere se o potravu) a holubice (rozdělí se o potravu). Kdo vyhraje? Mnoho jestřábů – nevýhodné pro všechny. Mnoho holubic – výhodné pro jestřáby. Zastaví se to na rovnovážné frekvenci. Mutant, který se chová jednou jako holubice, jindy jako jestřáb, vyhrává.
r-selekce - selekce k větší růstové rychlosti
K-selekce - selekce k větší konkurenceschopnosti
Názvy podle tradičního označení konstant v logistické rovnici.
dN/dt = rN(K – N)/K
N = počet jedinců v populaci, r = růstová rychlost, K = kapacita prostředí, odrážející maximální počet jedinců, které dané prostředí dokáže uživit
Náhodný výběr vede k selekci rychle se množících jedinců.
V určitých pozicích nukleotidového řetězce vznikají přednostně určité typy mutací.
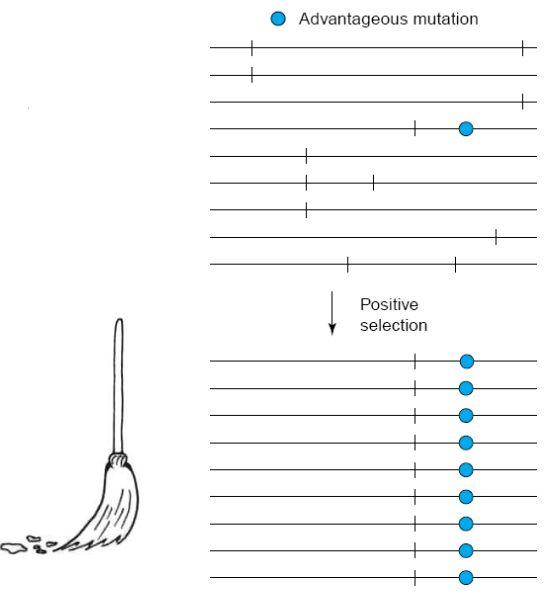
selective sweep
V okolí výhodné mutace dojde ke snížení genetické variability a zároveň ke zvýšení vazebné nerovnováhy v okolí výhodné mutace. Pokud je selekce výhodná, zafixuje se i blízké okolí.
evoluční svezení se, genetic hitchhiking
Evoluční osudy genů ležících na stejném chromozomu se vzájemně ovlivňují. Genetický draft je změna ve frekvenci alely v populaci díky genetické vazbě s jinou alelou (výhodnou či nevýhodnou)
Evoluční tahy jsou procesy probíhající na úrovni nosičů genetické informace. Působí společně s přirozeným výběrem a dalšími mechanismy.
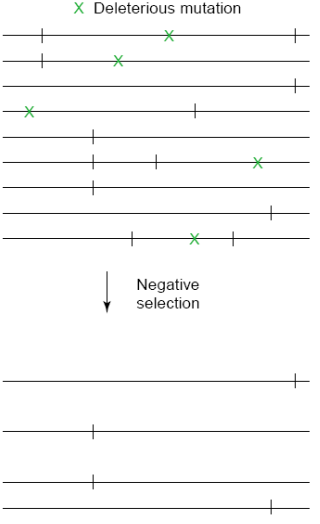
background selection
Dochází k odstranění alel z populace díky genetické vazbě s nevýhodnou mutací.
Vede ke snížení genetické variability, které však není tak výrazné jako v případě selekčního vymetení.
Některé alely se dokáží šířit na úkor jiných alel. Vnutí jim svou sekvenci, podle které se musí opravit. Jedná se o tzv. sobeckou DNA.
Patří sem procesy odpovědné za šíření jednotlivých variant repetitivních úseků DNA v rámci genomu a u pohlavně se rozmnožujících organismů i v rámci celého genofondu.
Př.
Při meiotickém tahu se do buněk přednostně dostává jedna alela a druhá se buď nedostává vůbec, nebo s menší frekvencí.
Jedna alela je přednostně vytlačena do pólové buňky alelou, která se dostala do vajíčka.
Při samičí meióze vzniknou 4 buňky, 1 dává vznik vajíčku a 3 pólové buňky
Kompenzuje mutační tah. Dochází ke vzniku mutací během reparačních procesů.
gametický tah
Výsledkem samičí meiózy jsou 4 spermie. Ze začátku jsou spermie propojené cytoplasmatickými můstky. Mohou si tak snadno do určité míry vyměňovat třeba mRNA.
Občas se stává, že dochází k likvidaci spermií, které nenesou upřednostňovanou alelu.
Př. t-haplotyp u myší
Mutační a reparační tah zodpovídají v největší míře za evoluci celkové struktury genomu, tedy za jeho zmenšování či zvětšování.